|
DMSTA
Calibration & prepared for U.S. Army Corps of Engineers & June 30, 2005 |
|
||||||||||||||||||||
|
Introduction
Datasets Calibration
Procedures Calibration
Results Model Testing
Discussion This page describes compilation of data and preliminary calibration of DMSTA for reservoirs. Subsequent refinements to the calibration are described at the DMSTA2 website. Current efforts to manage water quantity and quality in Florida (CERP, ACELER8, The Long-Term Plan) indicate that reservoirs are likely to become increasingly important components of regional water-management systems over the next decade or two. Forecasting tools are needed to support design and operation of reservoirs working in conjunction with other management measures (BMP's, STA's) to meet water quality standards while achieving other water management objectives. The Dynamic Model for Stormwater Treatment Areas (DMSTA) was developed to support design of wetland treatment areas to remove phosphorus from runoff and Lake Okeechobee discharges. The model has been calibrated to over 100 datasets derived from experimental tanks, field test cells, natural wetlands, operating STA's, wastewater treatment wetlands, and a few lakes north of the Okeechobee. Most of these datasets represented relatively shallow marshes dominated by emergent or submerged vegetation. This report describes model enhancements and calibration to support application to storage reservoirs. Hydrologic functions of reservoirs include capture of runoff and Lake regulatory releases, water supply, and flood control. Potential water quality benefits of reservoirs working in conjunction with STA's include:
The extent to which these benefits are realized will depend upon the designs of the reservoirs, STA's and conveyance systems, as well as upon how they are operated to achieve water quality and other water management objectives. The design model update allows simulation of interconnected reservoirs & STA's using the reservoir calibrations described below, as well as updated STA calibrations being developed separately. An interim report for this project describes proposed DMSTA enhancements and selection of CERP reservoirs for model demonstration and testing. The latter have been compiled from previous SFWMM simulations and other sources. They are used here for demonstration purposes and do not reflect current project designs. Simulation results can be viewed below. Previous evaluations of Restudy alternatives (Walker,1998) identified significant differences between reservoirs and STA's with respect to depth, hydraulic regime, and phosphorus removal performance, forecasted using steady-state models. Compared with typical marsh treatment areas and previous DMSTA calibration prototypes, initial CERP reservoir designs tend to have greater mean depths, greater variations in depth, and longer water residence times. Reservoirs in some regions north of Lake Okeechobee (e.g., Taylor Creek/Nubbin Slough) may also operate in higher concentration ranges (500-1000 ppb vs. 12 - 200 ppb for STA calibrations). These factors can be expected to have significant effects on vegetation communities, phosphorus dynamics, and model calibrations. Currently, STA's are operated at a static water depth of 1.2-1.5 ft. Deteriorations in vegetation integrity and performance have been observed in cells with prolonged water depths exceeding 2.5 to 3 ft (e.g., STA1W Cell 2). Current designs for CERP reservoirs have maximum depths ranging from 6 to 12 ft. Hydraulic and depth regimes are more predictable than vegetation characteristics and performance, especially given the high variance in reservoir depth that is not typical of the Florida lakes. While emergent marsh communities may develop in shallower peripheral regions or more extensively after extended periods of low water levels, reservoir plant communities are more likely to be dominated by suspended phytoplankton and possibly submersed vegetation (e.g. hydrilla). P removal may be inhibited to some extent by such factors as algal and non-turbidity (reducing light penetration and photosynthesis), wind mixing, and intermittent vertical stratification leading to oxygen depletion at the sediment/water interface and release of phosphorus from bottom sediments. Because of their greater depths, lack of internal levees, and lower hydraulic resistance, reservoirs are likely to be more susceptible than STA's to wind-driven waves and currents, which can discourage development of stable vegetation communities. These characteristics have been documented in Lake Okeechobee, which provides the largest but probably not the best prototype for the DMSTA reservoir calibration. DMSTA makes no attempt to model each of these processes directly. Their net effects contribute to variance in the model calibrations from one reservoir dataset to another. For this reason, the calibration datasets would ideally represent a wide range of depths, hydraulics, and vegetation likely to be characteristic of CERP reservoirs. The attached table summarizes datasets that have been compiled to support calibration of a phosphorus cycling model for reservoirs. Burns & McDonnell et al. (2004) conducted an exhaustive search of available lake and reservoir data from Florida. A inventory of available water quality data from over 600 water bodies with surface areas greater than 100 acres was compiled. Screening identified a short list of 36 lakes with potentially sufficient information. The final compilation included detailed data on 8 lakes and corresponding DMSTA input datasets. These have been combined with previous files from Lake Okeechobee and others north of the Lake. Specific sources include:
The regional distribution (excluding the EPA/NES lakes) is shown here. Two lakes (Harney & Istokpoga) appear in more than one source, each representing a different set of source data, time period, and/or data-reduction procedures. Based upon review of the source data, the calibration periods have been restricted to intervals with consistent inflow and outflow sampling and to allow for initialization of the phosphorus storage term in the model. While the collection of datasets overall provides a sufficient basis for model calibration, each dataset has limitations with respect to data quality and/or system characteristics that may reduce its usefulness as a calibration prototype for CERP reservoirs. Specific limitations include:
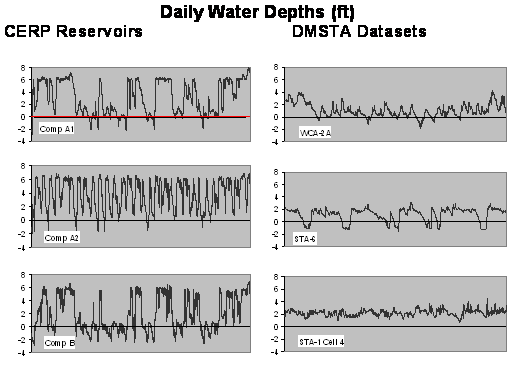
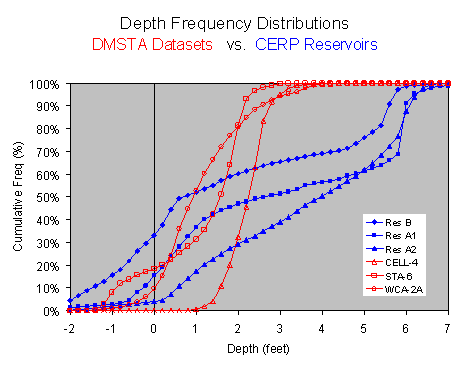
The attached figure compares depths, hydraulic loads, and inflow concentrations in the reservoir calibration datasets with typical reservoir & STA designs. While the datasets encompass the expected design ranges in mean depth, hydraulic load, and water residence time, there is little overlap. Four of the datasets (Istokpoga, Crescent, Jessup, Thonotosassa) have hydraulic loads within the reservoir design range. Mean depths are generally greater than the early CERP designs with 6 ft (183 cm) maximum depths (labeled A1, A2, B, C in the attached) but similar to the 12-ft (365 cm) design (AA) currently planned. The major limitation is that water depths are substantially more variable in the reservoirs, as indicated by depth ranges and time series. While one dataset (Poinsett) has minimum depths in the design range, it is not very useful for model calibration because its hydraulic load is far about the design range (40 m/yr vs. <20 m/yr) and simulations are insensitive to model parameters. The greater variations in depth reflects seasonal withdrawals for water supply that generally do not occur in the calibration lakes. The range of average inflow concentrations in the calibration datasets (49 - 1,100 ppb) covers the expected design range for reservoirs in and north of the EAA. Reservoirs in the urban areas (e.g., C11) will probably operate below this range. For example, inflow P concentrations to the C11-West Impoundment have been estimated at ~25 ppb (Mactec, 2004). Potential implications of these dataset limitations are discussed further below. Previous DMSTA calibrations were based upon a single prototype dataset for each vegetation category (Emergent, SAV, PSTA). Each prototype was selected based upon a variety of factors, including vegetation characteristics, surface area, duration of dataset, and monitoring intensity. The remaining datasets in each category were utilized for testing prototype calibrations. In this case, each reservoir dataset has strengths and limitations and no single one stands out as the "best" prototype. Accordingly, the following alternative procedure has been followed in calibrating and testing alternative models:
The procedure followed to initialize the simulations is similar to that applied to other datasets. The first 5-10 years of the time series is repeated twice and the calibration period begins 1-3 years after the start of the dataset. These intervals vary with dataset depending on the time needed for stabilization of P storage and the calibration time series. Further sensitivity analyses are performed to ensure that simulations are independent of the assumed initial storage. Averaging periods of 30-90 days are used in the simulations, depending upon dataset, monitoring frequency, and water residence time. From 1 to 4 stirred tanks are assumed, depending on lake shape. DMSTA's P cycling model includes a depth multiplier on the phosphorus uptake rate. Uptake rates are proportional to depth between 0 and 40 cm and constant thereafter for reservoirs. This multiplier was originally added to reduce correlations between model residuals and depth in marsh simulations. Because most mesocosm and steady-flow treatment wetland datasets do not indicate a depth dependence, this factor is thought to reflect a topographic effect; i.e., a decrease in effective surface area at shallow water levels because of deviations from the assumption of a uniform ground elevation and/or short-circuiting. Because of the higher depth regimes of the calibration datasets, there is no basis for determining whether this factor is appropriate for reservoir simulations based upon model residuals. Because a reduction in effective surface area or short-circuiting would also be expected to decrease P removal rates in reservoirs, the factor is included in each of the models tested below. This assumption does not influence the calibrations because depths in most of the calibration lakes remained well above 40 cm. A sensitivity analysis using reservoir design datasets indicates that average P removal rate would increase by ~1% in the reservoir with the most significant drawdown (Compartment C) if uptake rates were assumed to be independent of depth. Using the above procedures, three phosphorus cycling models have been calibrated and tested:
Simulations of each dataset using each model are attached. Model structures and results are described below. 1. First-Order Model The net P removal per unit area is assumed to be proportional to water-column concentration. The steady-state version of this model was first applied to European lakes by R. Vollenweider in the late 1960's and subsequently widely applied elsewhere. Walker & Havens (2003) applied it in dynamic form to Lake Istokpoga . It has one parameter (K1, m/yr) and no storage compartment (equivalent in DMSTA terms to C0 = 0 ppb and C1 = infinity). The reservoir is modeled essentially as a sedimentation basin removing particles with a fixed settling velocity. These particles may enter from the watershed or be generated in the reservoir in the form of phytoplankton, a fraction of which subsequently settles. Effects of re-suspension or recycling are embedded in the calibrated net settling rate. The median K1 value for these datasets (3.2 m/yr) is similar to that estimated by Walker & Havens (3.5 m/yr) for Istokpoga. Previous calibrations to Lake Okeechobee (Walker, 2000a ,2000b) had average K values declining from ~4 to ~1 m/yr between 1975 and1999, possibly as consequences of the loss of littoral zone, decrease in calcium content, and/or gradual accumulation of phosphorus in the "mud zone" or open lake sediments. The STA design model (Walker, 1995) has a similar P removal function but different mixing assumption (marsh sheet flow vs. well mixed) and vastly different calibrations (K1 = 10.2 m/yr for marsh vs. 3.2 m/yr for these lakes). The attached figure compares observed and predicted concentration reductions using the median K1 value calibrated to each dataset. The residual standard error is 15% for all lakes and 11% for lakes in the design hydraulic load range for CERP reservoirs (Istokpoga, Crescent, Thonotosassa, Jessup). 2. Second-Order Model 3. Biomachine Model The biomachine model
simulates the primary P fluxes between the water column and vegetation (uptake,
recycling, & burial). It was originally developed by R. Kadlec for
treatment wetlands and subsequently adapted for use in DMSTA. It's three parameters are defined relative to the steady-state
solution. These parameters and nominal calibrations for
reservoirs are listed below: In previous DMSTA calibrations to experimental mesocosms and marsh datasets, the parameters C0 and C1 were estimated independently from measured concentration and plant biomass phosphorus data. The parameter K was fit to concentration time series data using a non-linear parameter estimation routine that minimized the sums of squares of the log-transformed residuals (observed - predicted concentrations). Primarily because of the limited variance in these earlier datasets (many of which were steady-flow experimental mesocosms and test cells), the sensitivity to C0 and C1 was generally low and it was not possible to calibrate them directly to water column P time series. Recent calibration datasets for full-scale STA's and reservoirs have much greater variations in inflow volumes, loads, and water depths. In many cases, this increases sensitivity to C0 and/or C1 and enables least-squares calibration of two out of the three model parameters simultaneously. Direct measurements of biomass phosphorus are not available for the reservoir calibration datasets. This makes it impossible to calibrate C0 & C1 independently. Because of the higher concentration regimes in the reservoir datasets ( >40 ppb), sensitivity to C0 is generally low. Attempts at least-squares calibration of K and C0 simultaneously were largely unsuccessful because of the low C0 sensitivity and inverse correlation between the two parameters. Simultaneous calibration of K and C1 is feasible in some datasets with C0 constrained to 4 ppb, though confidence intervals for C1 are relatively wide. Joint confidence intervals for K & C1 indicate that lakes have substantially lower K values and higher C1 values, as compared with STA cells. One exception is Rodman Reservoir, which clearly clusters with the STA calibrations (see Discussion). The K/C1 calibration did not converge in all datasets, generally because of low sensitivity to C1 and/or long simulation periods required to initialize the P storage term at these low K values. The median C1 value is ~150 ppb (vs. 15-40 ppb for STA cells). To include all of the datasets in the joint calibration, a least-squares estimate of K has been derived for each dataset with C1 constrained to 150 ppb. The individual calibrations are shown in the attached table. The joint biomachine calibration is based upon median values for the individual datasets (K = 3.2 m/yr, C1=150 ppb). K values for the lakes in the design hydraulic load range vary from 2.3 to 5.9 m/yr. The residual standard error is 14% for all of the lakes and 9% for lakes in the design hydraulic load range. Results indicate that the biomachine model performs slightly better than the simpler first-order model without a storage term (RSE = 9% vs. 11% for lakes in design HLR range). The second order model is clearly inferior (RSE = 22%), which is not unexpected given that was originally developed from a nationwide dataset from reservoirs with greater depths and hydraulic loads, as compared with Florida lakes and typical CERP reservoir designs. The error distributions associated with the first-order and biomachine models are not distinguishable, given limitations of the datasets. For reasons discussed below, the biomachine calibration is recommended for use in reservoir design. Reservoir simulations indicate that predicted P removal rates are relatively insensitive to the choice of model. The biomachine calibration is further described and tested in the attached figures and tables . Observed and predicted outflow concentrations an loads are compared. For all datasets, the model explains 88% of the variance in the flow-weighted-mean outflow concentrations and 61% of the variance in the percent reductions in concentration. Residual standard errors are similar in magnitude to the DMSTA marsh community calibrations. Correlations between calibrated K values and dataset features provide further tests of model generality. The spread of K values in the low hydraulic load range (<20 m/yr) is 1.8 - 5.9 m/yr (excluding the Okeechobee 1995-1999 calibration, 1.2 m/yr). This is roughly a 2-fold variation around the joint calibration of 3.2 m/yr. This spread can be used to estimate the uncertainty associated with model forecasts (see reservoir simulations). As expected, variance in K increases at higher hydraulic loads as simulation sensitivity to K decreases and sensitivity to random errors in the inflow/outflow concentration increases. Model residuals (observed - predicted concentrations, log transformed) are analyzed to identify the correlations with season, length of dataset, and lake features (depth, hydraulic load, inflow concentration, etc). This analysis has been applied to dynamic residuals (quarterly or monthly flow-weighted means from all datasets combined), yearly flow-weighted means, and to long-term flow-weighted-means. Residuals appear to be independent of season.. Yearly residuals are uncorrelated with depth (80 - 300 cm), hydraulic load (0.5 - 70 m/yr), and outflow P concentration (40-1000 ppb). These ranges define the limits of model applicability. The model tends to under-predict outflow concentrations at depths > 300 cm ( see Discussion) Further testing of the biomachine and first-order calibrations is based upon data collected by the EPA's National Eutrophication Survey (NES) in 1973. The NES provided average water and phosphorus balances based upon a single year of monitoring (vs. 3-16 years for the DMSTA calibration datasets). If the P balance of each lake is assumed to be at steady-state, the data can be used to test the steady-state versions of the DMSTA calibration. Deviations from that assumption are reflected in the error distributions. Some of the NES lakes are also included in the DMSTA calibration datasets (Okeechobee, Jessup, Istokpoga, George, & Crescent). Results are expected to vary as a consequence of difference in time frame (1973 vs. 1978-2003) and duration of monitoring (one year vs. multi-year). Calibration of the steady-state model (computation of K) based upon a single year of data is subject to error because it ignores changes in phosphorus storage during the year, as estimated by the dynamic simulations, can be substantial. This effect is expected to be smaller in the long-term term DMSTA datasets. The steady-state solution of the biomachine and first-order models can be approximated by R = K' / ( K' + Q ), where R = percent reduction in concentration, K' = settling rate for steady-state model (m/yr), and Q = hydraulic load (m/yr). This is an approximation because it does not account for differences in mixing regime (stirred tanks) or C0 effects. When applied to the DMSTA calibration lakes with a K' value of 2.8 m/yr, the steady-state model predicts average concentration reductions that are similar to DMSTA simulations using the biomachine calibration (K = 3.2 m/yr, C1 = 150 ppb, C0 = 4 ppb). The difference between K' and K reflects the simplifying assumptions and the flow/concentration dynamics that are not accounted for in the steady-state model. Observed and predicted concentration reductions for each dataset are shown in the attached figure. Predictions are shown for 2-fold variations of K' around the calibrated value of 2.8 m/yr (1.4 - 5.6 m/yr). Some of the NES lakes were heavily impacted by point sources. Lakes with inflow P concentrations exceeding 500 ppb are excluded. The standard error of the predicted concentration reductions is 12% for the calibration lakes and 23% for the NES lakes. A higher residual variance is expected because the NES data are one-year snapshots of the lake P balances. Several lakes have concentration reductions exceeding the calibration envelope. With the exception of Rodman, all of these lakes have predicted outflow concentrations greater than 100 ppb. Marion, Mineola, and Weohyakapka had outflow concentrations in the 23-30 ppb range, as compared with the 40 ppb lower bound for the DMSTA calibration lakes. Two of these are at the lower edge of the calibration envelope and the other is in the middle. Compilation of modern data from these lakes may support further testing of DMSTA calibrations in the lower concentration range. Despite
the higher residual variance, DMSTA calibrations are consistent
with data from most of the NES lakes and conservative for others. The
steady-state version should not be applied to reservoir designs, which are
likely to have greater variance in depth and concentration, as compared with the
calibration lakes. This would tend to increase the difference between K' and K.
Given the limitations of the datasets and calibration result, it is difficult to distinguish the performance of the first-order and biomachine models. Several other methods for fitting and testing the models have been explored and found to give similar conclusions. While the first-order model is attractively simple, biomachine K/C1 calibrations indicate that inclusion of storage/recycling components improves simulations of the outflow concentration dynamics in some of the datasets. If storage is primarily in the form of labile inorganic P in bottom sediments (vs. vegetation), other storage/recycling models that do not compute the uptake rate as a function of storage may be more appropriate. Net P settling rate is highly correlated with sediment accretion rates in midwestern lakes & reservoirs (Fig 3). Estimates of sediment accretion rate are typically required to drive coupled water column/sediment mass balance models (Walker, 2000a Fig 5 , 2000b Fig 15, 2001 Fig 11, 2003 Fig 9 ). Future enhancements of DMSTA to include a-biotic storage compartments (floc and/or soil) will provide a framework for testing alternative reservoir models. The existing database does not support addition of these additional compartments or other model refinements , however, especially given the limited depth ranges relative to the CERP reservoir designs and other dataset limitations. While the datasets provide a sufficient basis for model calibration to support reservoir performance forecasts, it is possible that the larger database compiled by Burns & McDonnell (36 lakes) contains lakes with more representative depth, hydraulic load, and/or concentration regimes. Further investigation is recommended. Finding lakes that have both sufficient data and highly managed water levels may be difficult, however. Eventually, monitoring and simulation of pilot-scale or full-scale reservoirs will provide the best basis for refining model structure and/or calibration. Rodman Reservoir has a high K value relative to the median calibration (21 vs. 3.2 m/yr). It clearly clusters with STA cells in the K/C1 calibrations. This suggests that Rodman has unique features that promote phosphorus removal. It's relatively high length/width ratio stands out, but simulations attempt to account for this with 4 stirred tanks. The aspect ratio may limit wind mixing, a factor which is not considered in the model. The performance may be related to unique vegetation characteristics (SAV). Further investigation is recommended. A vegetation inventory for all of the calibration datasets may help to explain some of the variations in the calibration results. This information may also help to develop guidance for reservoir design and/or vegetation management to promote phosphorus removal. Model residuals suggest that outflow concentrations are under-predicted at depths > 300 cm or 10 ft. These data are from Lakes Harney and Okeechobee. Lake George (not reflected in the residuals plots) is also relatively deep and has a negative P removal efficiency. The pattern in the residuals may be related to depth or to other factors associated with these lakes. The model may under-predict the outflow concentrations of the deeper reservoirs currently being contemplated (max depth 12 ft). Risk of light limitation and of vertical stratification leading to phosphorus release from anoxic sediments both increase with depth. The data are insufficient to support design recommendations. It may be possible to modify reservoir operating depths if monitoring data indicate that vertical stratification is a significant factor hindering P removal. See Reservoir Simulations for further discussion of these preliminary simulations. Further analysis of the reservoir data and refinements to the calibration are described at the DMSTA2 website. Everglades Restoration Efforts: CERP ACELER8 Long-Term Plan Water Quality Evaluation of Restudy Alternatives - 1998 Potential DMSTA/EPGM Applications to CERP Projects - March 2003 CERP DMSTA Workshop - October 2003 DMSTA Overview - R Kadlec - GEER Conference 2003 DMSTA Home
Current DMSTA Calibrations
DMSTA Research Datasets
North of Lake Okeechobee
Datasets |
|||||||||||||||||||||
|
http://www.wwwalker.net/dmsta/reservoirs/index.htm 07/08/2005 |
|||||||||||||||||||||

{kind=link}
{kind=link}